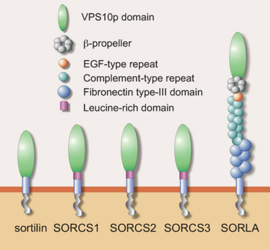
The sortilin family contains five members, Sortilin, SorLA and SorCS1-3, that are all particularly expressed in developing and adult neuronal tissue. Their common characteristic, the hallmark of the family, is the luminal Vps10p-domain named after the yeast sorting protein Vps10p. This domain is not found in other mammalian proteins and is remarkable in the sense that it binds a number of ligands, notably neurotrophic growth factors and peptides. The latter include (pro)NGF, (pro)BDNF, GDNF and neurotensin, factors that are needed for the differentiation of nerve cells and to sustain normal function of the nervous system. The C-terminal cytosolic part of the sortilins contains sequence signals for binding to various sorting adaptor proteins.
PROCESSING
The Sortilins are subjects to a number of posttranslational modifications. Apart from glycosylation, this in particular concerns enzymatic cleavage aimed at specific N- and C-terminal sites in the extracellular part of the receptors. Three types of functional cleavage are known. One mediates release of a short N-terminal propeptide from each receptor, another conveys shedding of the entire ligand-binding ectodomains, and a third converts one of the family members from a single to a two-chain receptor.
Propeptide-cleavage: In all five receptors the Vps10p-domain contains a short N-terminal propeptide which is removed by cleavage during or after passage of the TGN. The propeptide may have at least two functions. In the first place it prevents binding of ligands to the Vps10p-domain of Sortilin and SorLA, which can bind their own propeptide. Accordingly, it appears that the propeptides of these receptors occupy the ligand-binding central tunnel of the Vps10p-D b-propeller. In contrast, SorCS1-3 do not bind their respective propeptides and do not depend on propeptide cleavage for ligand binding.
Another and separate function of the propeptide is to facilitate processing of newly synthesized receptors. The expression of SorLA and SorCS3 does not depend on the propeptide, whereas that of Sortilin, SorCS1 and SorCS2 is seriously hampered in the absence of their respective propeptides. Thus, in the latter three receptors the propeptide may act as an intrinsic chaperone and prevent minor misfoldings hampering processing and transport in the proximal biosynthetic pathway.
Shedding: Similar to many other type 1 receptors the Sortilins may shed their luminal domains. Sortilin and SorCS2 are the least affected and show only minor shedding (<0.5% per hour) while 20-40% of SorCS3 and SorCS1, and about 4% of SorLA, are shed from CHO-transfectants within one hour. Various findings suggest that differential enzymatic activity may provide a cell dependent regulation of shedding. Yet, TACE/ADAM17 seems to be the main responsible proteinase and with the exception of SorCS2, ectodomain shedding is not observed in TACE-deficient CHO-cells. Accordingly, release of ectodomains is markedly increased by stimulators of metalloprotease activity, e.g. PMA and PDGF-BB, but unlike the ectodomains of Notch and p75, shedding of the Sortilins do not relate to ligand binding or expression on the surface membrane. Following cleavage of the ectodomains at least SorCS1 and SorLA are targeted by g-secretases and release their cytosolic domains. The functional implications of Vps10p-D receptor shedding are still unclarified . Yet, the shed ectodomains may complex with ligands and serve to regulate the activity of ligands in solution and it is an interesting possibility that the subsequent release of cytoplasmic domains may participate in intracellular events, e.g. signalling, prior to their degradation in proteasomes.
A third and completely different type of ectodomain cleavage concerns SorCS2, which unlike any of its relatives, is converted from a single to a two chain receptor by enzymatic processing. If and how this impacts on receptor function remains to be seen.
<strong>TRAFFICKING
The cytoplasmic domains of Sortilin and SorLA contain several sorting-motifs and exhibit functional interaction with several cytosolic adaptors including AP-1 and -2, GGA1-3, and elements of the retromer complex. Accordingly both receptors are capable of rapid endocytosis and exhibit vivid intracellular trafficking notably between endosomes and the trans-Golgi network (TGN). Only about 10% of the receptors are found on the plasma membrane and it therefore seems likely that the two are mainly engaged in intracellular protein transport and sorting. Their targets and specific functions in this context, however, are only beginning to emerge.
Unlike Sortilin and SorLA, SorCS1-3 are not high performers in terms of trafficking. Both SorCS2 and -3 are mainly expressed on the cell surface and contain functional tyrosine based motifs for endocytosis, but neither of the two exhibits major intracellular trafficking. SorCS1 is more complicated due to the existence of several isotypes (a, b, and c in humans) which have identical luminal domains but different cytoplasmic tails. SorCS1a is mainly intracellular, type-b is almost only found on the plasma membrane, whereas type-c is equally distributed between the cell surface and intracellular compartments. Accordingly, only isotypes a and c contain functional motifs for endocytosis. Also, vesicular SorCS1 does not appear to partake in Golgi-endosome transport. It is interesting though that SorCS1-a and (in particular) -b contain one or more SH2/SH3-domain binding motifs. Considering that SorCS1 is a major target for shedding and subsequent g-secretase activity, it could be speculated that the cytoplasmic domain(s) plays an independent role e.g. in signalling.